
Common Dolphin

Created July 2023
The common dolphin is likely the most abundant and widespread species in the world, it occurs in tropical and temperate coastal and pelagic marine waters worldwide. Common dolphins have a lean body and are easily to identify by their pale-yellow patch on the anterior and the very unique criss-cross pattern on their sides, which distinguishes them easily from similarly shaped dolphins This very social species is found in groups of tens to thousands of animals, that are fast swimmers and like acrobatics, and schools are being usually very active.
The species is not a resident but a visitor in the waters of NAMMCO member countries.

Abundance
The global population is about six million, with 640.000 common dolphins in the North Atlantic (Hammond, 2017; Hayes, 2022).
Distribution
Common dolphins are found worldwide in warm-temperate and tropical waters, generally between 60°N and 50°S. Their distribution varies depending on the season and sea-surface temperatures.
Relation to humans
The species is hunted for human consumption or shark bait in certain parts of the world, but not in the North Atlantic. By-catch in fishery is, however, very common.
Conservation and management
The common dolphin is a widespread and abundant species. As a result, the species is listed as “Least Concern” on both global and regional European Red List in the most recent assessment (2021 and 2023, respectively) of the IUCN.
Meanwhile, by-catch is the main threat that common dolphins face today, with 1000s being by-caught every year off Europe, with a hot spot in the Bay of Biscay.
The common dolphin is protected in the North Atlantic. It is only a visitor in the waters of NAMMCO member countries, therefore no assessment effort has been dedicated to this species.

Scientific name: Delphinus delphis
Faroese: Springari
Greenlandic: not given
Icelandic: Léttir
Norwegian: Gulflankedelfin
Danish: Almindelig delfin
English: Common dolphin
Lifespan
The maximum lifespan for common dolphins (for both sexes) is estimated at 30 years.
Size
Adults reach between 150 and 220 cm in length and can weigh up to 235 kg.
Productivity
Females give birth to a calf every one to four years after a gestation period of 10 to 11.7 months.
Movements
Common dolphins do not undertake large migrations but rather tend to stay in areas where prey is abundant.
Feeding
At greater depths, common dolphin prey on squids and small fish such as scombroids and clupeoids, but they also hunt for schooling fish (such as herring, sardine, mackerel or smelt) and crustaceans (such as pelagic red crabs and krill) at shallower depths.
General Characteristics
Common dolphins are, at the present time, among the most abundant cetaceans worldwide. They have a lean body with a relatively long, delimited beak and high, moderately curved dorsal fin. Additionally, a criss-cross colour pattern extends across their whole body, crossing mid-body underneath the dorsal fin. This pattern results, in detail, in a four-part pattern of dark grey to black coloration on the upper side of the body, a white underside, a pale yellow patch on the anterior and grey posterior patch (Perrin, 2018).
The criss-cross pattern on the common dolphin’s sides distinguishes them from similarly shaped dolphins such as the spinner and striped dolphin as well as the similarly coloured (but with narrow stripes) Atlantic white-sided dolphin (Carwardine 2020).
Common dolphins are known for bow riding the waves created by boats, large cetaceans and even basking sharks in particular (Massett et al., 2020; Würsig, 2018).
Taxonomy
Common dolphins belong to the Delphinidae family and Delphinus genus. The genus used to consist of two species (short-beaked and long-beaked common dolphins, D. delphis and D. capensis respectively), but more recent research questions this split and since 2016 the common dolphin has been considered a single species (Natoli et al., 2008; Perrin, 2018; Pinela et al., 2011). At this point, four subspecies are recognised: the short-beaked common dolphin (Delphinus delphis delphis) which is the nominate subspecies, the Eastern North Pacific long-beaked common dolphin (D. d. bairdii), the Indo-Pacific common dolphin (D. d. tropicalis) and the Black Sea common dolphin (D. d. ponticus) (Perrin, 2018).
Did you know?
The common dolphin (Delphinus delphis) has an unexpected counterpart, the common dolphinfish (Coryphaena hippurus)

Life History and Ecology
Adults reach lengths between 150-225 cm for both sexes and can weigh up to 235 kg (Murphy et al. 2006a, Perrin 2018). The maximum estimated age is 30 years (in the western North Atlantic). Sexual maturity varies with regions but is estimated at 3 years for the Black Sea population, 7-12 years in the eastern Pacific and Western Atlantic for males. Females are thought to reach sexual maturity between 2-4 years in the Black Sea region while 6-8 years in the eastern Pacific and western Atlantic region (Perrin 2018). The mating system between common dolphins seems to be promiscuous dependent on sperm competition (Murphy et al. 2005).
Females are estimated to be pregnant for 10-11.7 months before giving birth to a calve between 80 and 93 cm in length (Murphy et al. 2006a, 2009, Perrin & Reilly 1984). The calving interval varies per region from about 1 year in the Black Sea region compared to about 4 years in the eastern Atlantic (Perrin 2018) .
Social interactions
Common dolphins are evidently a very social species. For instance, schools of great variety (10 up to 10,000) can occur which often split in smaller subunits of 20-30 individuals (Evans 1994). Moreover, they are forming fission–fusion dynamics due to the occurrence of highly fluid societies driven by prey availability, predation risk, age, sex, and reproductive status (Ball et al. 2017, Castro et al., 2022, Young & Cockcroft 1994, Zanardo et al. 2016).
Group composition and sizes likely vary seasonally, but mixed groups of males and females with sub-adults and calves are common (Carwardine 2020). At this time, genetic analyses suggest that the members of a school are not necessarily closely related, and there may be group segregations depending on age and sex (Ball et al. 2017, Viricel et al. 2008).
Common dolphins occur regularly in association with other species such as pilot whales (Globicephala spp.), striped dolphins (Stenella coeruleoalba), several oceanic dolphins (Lagenorhynchus spp.) and Risso’s dolphins (Grampus griseus) (Evans 1994, Frantzis & Herzing 2002).
Sound/Communication
As a highly vocal species, the common dolphin repertoire includes clicks, whistles, burst pulse calls and buzzes (Moore & Ridgway 1995). Their clicks are very short, high frequency sounds (23 to 100 kHz) in order to sense their environment and locate prey (Moore & Ridgway 1995, Soldevilla et al. 2008). Burst pulses are a series of rapidly produced clicks (up to 10 times faster than echolocation clicking) and are a communication tool. Moreover, buzzes, a rapid succession of clicks that seem to be a continuous sound to the human ear, and whistles; most commonly simple up- and downsweeps between 3kHz and 24kHz, are also used to communicate (Carwardine 2020). Like many other delphinid species, common dolphins produce unique, so-called ‘signature whistle’ that last between 1-4 seconds and serve as a name tag for each dolphin (Fearey et al. 2019).
Listen to common dolphin sounds (© NEFSC-NOAA) here:
DID YOU KNOW?
A common dolphin was communicating in “porpoise”
Within a restricted area in the Firth of Clyde in Scotland, a solitary common dolphin has been exposed to harbour porpoise vocalisations and observed interacting with them for at least 14 years. Interactions with other common dolphins have not been observed during that time.
The two species however communicate quite differently. Harbour porpoises mainly produce high-frequency clicks while common dolphins have an array of sounds from clicks, to whistles and burst calls in lower frequencies. Now, it seems like the common dolphin is “porpoising” and produces clicks with temporal and spectral characteristics that resemble porpoise clicks while stopping other vocalisations.
Read the study by Cosentino et al. (2022) here
Swimming and Diving Behaviour
Common dolphins exhibit aerial behavior, like many other dolphin species. For example, they leap out of the water to perform acrobatics, somersaults, and slapping the water surface with their flippers or lobtailing. The leaps can reach heights of up to 6-7 m. A common aerial display for example is called a ‘pitch-pole’ in which the dolphin aims to produce the largest splash after jumping straight up from the surface. Moreover, common dolphins are fast swimmers, with speeds reaching up to 40km/h when porpoising (Carwardine, 2020).
Dives up to 280 m have been recorded for as much as 8 minutes, though most foraging dives are much shallower and remain around 50m of depth that last between 10 seconds and 3 minutes (Perrin, 2018; Carwardine 2020).
Food and Feeding
The diet of common dolphins varies as much as the range of habitats they reside in and varies with season and region (Ambrose et al., 2013; Evans, 1994). At greater depths, common dolphin prey on squids and small fish such as scombroids and clupeoids, but they also hunt for schooling fish (such as herring, sardine, mackerel or smelt) and crustaceans (such as pelagic red crabs and krill) at shallower depths (Carwardine, 2020; Marçalo et al., 2018; Perrin, 2018). In some regions, common dolphins tend to feed during the night as their prey that usually occurs in deeper ocean layers migrates towards the shallower depths during the night (Henderson et al., 2012; Simonis et al., 2017).
In another case, stomach content from stranded animals from the Bay of Biscay, the Irish coast and bycaught animals in the Irish tuna driftnet fishery showed that oceanic short-beaked common dolphins’ diet generally consisted of fish, predominantly myctophids (the family of lanternfish) (Brophy et al., 2009; Pusineri et al., 2007).


Predators
Natural predators of common dolphins include killer whales and sharks (Biton Porsmoguer et al., 2015) while pilot whales and pygmy killer whales might also attack them (Carwardine, 2020).
Moreover, common bottlenose dolphins (Tursiops truncatus) have attacked and killed a short-beaked common dolphin in Ria de Arousa (Northwest Spain), presumably due to competition for food or practice for infanticide (Methion & Díaz López, 2021).
Health – Diseases and Parasites
Like all other cetaceans, common dolphins can be affected by various diseases and may be infected by parasites.
Diseases
The morbillivirus is one apparent disease in common dolphins, which also occurs in many other dolphin species as well. For example, it was found in six of 18 common dolphins that stranded along beaches of southern California (Reidarson et al., 1998) and in stranded animals in the Black Sea (Birkun et al., 1999).
Different species of the aquatic pathogen Streptococcus sp. have been found in stranded common dolphins. For example, in Australia, abscesses, sepsis and mortality were associated with Streptococcus iniae in a free-ranging short-beaked juvenile common dolphin (Souter et al., 2021). Furthermore, bacterial presence of Streptococcus agalactiae occured in urine, lungs and pericardial fluid in a stranded, wild male common dolphin that was found dead in September 2020 on the seashore next to the city of Bat-Yam, Israel (Morick et al., 2022).
Parasites
Internally, dolphins can host various types of parasites. For example, the nematode Anisakis sp. is a common parasite in the guts of various cetaceans and is present in common dolphins as well. Overall, Anisakis infections can leave ulcerative lesions in the stomach and, in severe cases, lead to haemorrhage and stomach perforation which can result in the death of the animal (Jaber et al., 2006).
When comparing the severity of Anisakis infections in common dolphins which stranded along the north-west Spanish coast in 1991-1996 and 2017-2018, Pons-Bordas et al. (2020) found a strong increase of infected dolphins and abundance of ulcerative lesions per host.
In another study on stranded common dolphins on the north-west Spanish coast, the protozoan parasites Giardia duodenalis and Cryptosporidium parvum were found in the animals (Reboredo-Fernández et al., 2014). The authors do not identify the parasites as the reason for the stranding, however, these results indicate the contamination of the marine environment by zoonotic pathogens.
Strandings
Several mass strandings of common dolphins have been widely reported in the North Atlantic, for example:
In 2002, 100 dolphins stranded in a shallow cove in France after probably being trapped during feeding at low tide (Dabin et al., 2002 in Viricel et al., 2008).
In 2008, 26 common dolphins stranded in Cornwall, UK, a suspected cause were naval exercises which took place four days before the event (Jepson et al., 2013).
Between the 12th of January and 16th of February in 2012, 178 common dolphins stranded in Massachusetts, USA (NOAA Fisheries, 2012).
Distribution and Habitat
Common dolphins inhabit tropical to temperate waters globally, with a preferred sea surface temperature of 10-28°C extending from about 40°N in the North Pacific and 60°N in the North Atlantic to 50°S. Likely due to the water preference, they may at times occur outside the normal distribution range when following warm-water currents (Carwardine, 2020). In some regions, populations are distinct to the extent of being considered sub-species (for instance the Black Sea population).
Within their range, they can be found anywhere between near-shore waters to several thousand kilometres offshore, although most common dolphin populations show a preference for areas with a seamount which are known for their strong upwelling (Amaral et al., 2012; Bearzi et al., 2003; Perrin, 2018).

Common dolphins are only visitors in the waters of NAMMCO member countries and have been rarely observed during the North Atlantic Sightings Surveys (NASS series 1987-2016, NAMMCO Scientific Publications, 2019). They have been more often observed however when surveys were conducted north of about 40°N like in 1987 when Spain participated, and in 1995, when the Faroe Islands surveyed west of the British Isles, see map below (Cañadas et al., 2009).

On another note, common dolphins have been observed many times during the SCANS Surveys and the ObSERVE survey west of Ireland (see maps).


Movement and migration
Common dolphins do not undertake large migrations but rather tend to stay in areas where prey is abundant. However, seasonal offshore movement during summer months of short-beaked common dolphins in the Northeast (NE) Atlantic may be linked to prey movement and the energetic demands of pregnant and lactating females (Brophy et al., 2009). Moreover, seasonal movements linked to prey and pregnant and lactating females have also been observed in South Africa (Cockcroft & Peddemorst, 1990).
Stocks and Abundance
Worldwide
Common dolphins are among the most abundant cetaceans worldwide, with a global population estimated to about six million (Hammond et al., 2008). Meanwhile the available abundance estimates are primarily from the Northern Hemisphere. Although the Southern Hemisphere constitutes a large portion of the species range, few abundance estimates are available (Braulik et al, 2021).
North Atlantic
Northeast Atlantic
In the NE Atlantic, common dolphins appear to form one large panmictic population based on genetic analyses (Moura et al., 2013) while between the central‐east and the NE Atlantic research reports a marginal level of differentiation (Natoli et al., 2006).
In the NE Atlantic, several surveys have been carried out for different small cetacean species, including short-beaked common dolphins. One example is the multinational Small Cetacean Abundance in the North Sea and Adjacent Waters (SCANS) surveys, a series of surveys that took place in 1994, 2005, 2016 and 2022.
The abundance for 2016 was estimated to be 441,500 (CV = 0.305) (Hammond et al., 2021). However, this estimate did not include contemporaneous aerial survey data from the Irish Exclusive Economic Zone which reported 33,215 (CV = 41.52; 95% CI: 19,844–55,595) possible common dolphins in this area (Rogan et al., 2018).
Northwest Atlantic
A separate genetically and morphologically distinct population exists in the Northwest Atlantic (Mirimin et al., 2009; Natoli et al., 2006). Due to the relatively low observed level of genetic differentiation across the whole North Atlantic a recent population split or high level of gene flow between two or more populations is hypothesized (Mirimin et al., 2009).
In the Northwest Atlantic the current abundance estimate is 172,947 (CV=0.21) with a minimum estimate of 145,216 (Hayes et al., 2022).
NAMMCO waters
The common dolphin is only a visitor in the waters of NAMMCO member countries. It is protected by all NAMMCO member countries.
No assessment effort has been dedicated to this species within the NAMMCO framework.
Globally
The common dolphin is a widespread and abundant species. The global population is listed on the IUCN Red List as “Least Concern” on a global level (Braulik et al., 2021), although the Mediterranean population is listed as “Endangered” (Bearzi, 2012).
The Convention on the Conservation of Migratory Species of Wild Animals (CMS) lists common dolphins in Appendix I and II. These conventions in general provide for international cooperation in the management of species like the common dolphin that are widely distributed.
All cetaceans, including the common dolphin, are listed as species in need of strict protection in Annex IV in the European Habitats Directive (European Directive on the Conservation of Natural Habitats and Wild Fauna and Flora, 92/43/EEC). It is an environmental legislation, which carries a financial penalty if member states fail to implement its requirements. In general, the overarching aim of the Habitats Directive is to achieve favourable conservation status for the species and habitats listed.
The Agreement on the Conservation of Small Cetaceans of the Baltic, Northeast Atlantic, Irish and North Seas (ASCOBANS) as well as the Agreement on the Conservation of Cetaceans in the Black Sea, Mediterranean Sea and Contiguous Atlantic Area (ACCOBAMS) cover the species.
Human Impacts
Hunting
Common dolphins are not subject to direct hunt in the NE Atlantic at the present time. In the past however, fishermen sometimes harpooned them as a source of fresh meat onboard.
Common dolphins were hunted historically by several countries until at least the 1990s, with over 1.5 million individuals taken between 1931 and 1966 in the Black Sea alone prior to a ban of commercial hunting of small cetacean species (Zemsky, 1996).
Today, direct take is still practiced in some areas including Japan, Taiwan, Venezuela, Peru and Mexico for consumption as well as bait (Carwardine, 2020; Kasuya & Perrin, 2017; Mintzer et al., 2018).
By-catch
Trawl, gillnet and purse-seine fisheries are threats to common dolphins globally, although the precise impact of by-catch on the populations is not certain (Murphy et al., 2006b; Thompson et al., 2013). Moreover, in European waters, common dolphins together with harbour porpoises (Phocoena phocoena) are the most vulnerable species to bycatch (Hammond et al., 2013). For instance, the ICES Working Group on Marine Mammal Ecology identified by-catch as the greatest threat for common dolphins in the Celtic Seas and Bay of Biscay/Iberian Peninsula (ICES, 2015).
Since 2016, strandings of small cetaceans with evidence of bycatch occurred in unprecedented numbers along the Bay of Biscay coast, breaking annual records (Dars et al., 2019). In addition, between 5,000 and 10,000 bycaught common dolphins were estimated in French Atlantic waters each year from 2016 to 2018 (ICES 2020).
Prey depletion
Over-fishing of prey species is more of a threat to common dolphins in the Mediterranean than in the NE Atlantic (Murphy et al., 2021). It has been observed in Spain (Cañadas & Hammond, 2008), Italy (Mussi et al., 2021) and Greece (Piroddi et al., 2011) in particular.
Noise/Disturbance
Human-made noise is a threat to most cetaceans, including common dolphins. For instance, military activity involving sonar can be dangerous to marine mammals and is a possible cause for a mass stranding of common dolphins that occurred in the UK in 2008 (Jepson et al., 2013 – see above under 3 – Life History and Ecology). In another case, underwater explosions from construction or other sources are further threats to this species (Danil & St. Leger, 2011).
Plastics
In the stomachs of 35 common dolphins, stranded between 2005 and 2010 on the Galician Coast, Spain, an average of 12 microplastic items (MP, < 5 mm in size) were found.
In detail, Dool & Bosker (2022) estimate that common dolphins ingest 76 MPs/day in the NE Atlantic based on a systematic review of 58 papers.
Several hundreds of chemicals, toxic heavy metals, pesticides, and pathogens are either added during manufacturing or adsorbed by MPs from the environment. Therefore, they become vectors of pollutants and toxins, which, along with the biomagnification and bioaccumulation process, make MPs a potential threat to the well-being of aquatic mammals (Nabi et al., 2022 ). Possible threats due to high exposure to MPs, are a higher risk of cancer and the exposure to accumulated pathogens on the MPs, but no studies are available linking MPs directly with cancer in marine mammals or showing that pathogen-colonized MPs can transfer pathogens (Nabi et al., 2022).
Nelms et al. (2019) found a possible relationship between the cause of death category and microplastic abundance, indicating that animals that died due to infectious diseases had a slightly higher number of particles than those that died of trauma and other drivers of mortality. It was not possible, however, to draw any firm conclusions on the potential biological significance of this observation.
Contaminants
As well as other marine mammals, common dolphins ingest persistent organic pollutants (POPs) by consuming fish that have been exposed to these compounds from coastal runoff and human industrial activities.
Concentrations of polychlorinated biphenyls (PCBs) in blubber of female common dolphins from the Atlantic coast of Europe were in 40% of all cases frequently above the threshold at which effects on reproduction could be expected (Pierce et al., 2008). Additionally, a few individuals of the common dolphins sampled had died from disease or parasitic infection. In the final analysis, persistent organic pollutants profiles in common dolphin blubber were related to individual feeding history.
Climate change
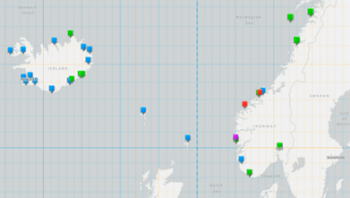
Strandings records from 1984-2018. Green=short-beaked common dolphin, blue=striped dolphin, red=Risso’s dolphin (Hansen, 2020)
Climate change will certainly affect cetacean species in various ways according to their distribution and temperature requirements. For some there may be positive impacts (e.g. a widening of the possible geographical distribution) while for others it will be negative (particularly Arctic species for which suitable habitat can shrink due to rising temperatures and loss of sea ice). The consequences for the common dolphin are not clear, but the changes in ocean temperatures may extend their distribution northwards. An indicator for this are increased observations of common dolphin strandings in Iceland and Norway since 1997 (see map, Hansen, 2020)
As common dolphins are only rare visitors in the water of the NAMMCO countries, there has not been conducted any specific research projects. However, sightings of common dolphins are recorded during the NASS surveys organised under the NAMMCO framework.
References
Amaral, A. R., Beheregaray, L. B., Bilgmann, K., Boutov, D., Freitas, L., Robertson, K. M., Sequeira, M., Stockin, K. A., Coelho, M. M., & Möller, L. M. (2012). Seascape Genetics of a Globally Distributed, Highly Mobile Marine Mammal: The Short-Beaked Common Dolphin (Genus Delphinus). PLoS ONE, 7(2), e31482. https://doi.org/10.1371/journal.pone.0031482
Ambrose, S. T., Froneman, P. W., Smale, M. J., Cliff, G., & Plön, S. (2013). Winter diet shift of long-beaked common dolphins (Delphinus capensis) feeding in the sardine run off KwaZulu-Natal, South Africa. Marine Biology, 160(7), 1543–1561. https://doi.org/10.1007/s00227-013-2208-6
Ball, L., Shreves, K., Pilot, M., & Moura, A. E. (2017). Temporal and geographic patterns of kinship structure in common dolphins (Delphinus delphis) suggest site fidelity and female-biased long-distance dispersal. Behavioral Ecology and Sociobiology, 71(8). https://doi.org/10.1007/s00265-017-2351-z
Bearzi, G. (2012). Delphinus delphis. The IUCN Red List of Threatened Species 2012: e.T134817215A195829089. https://doi.org/10.2305/IUCN.UK.2012
Bearzi, G., Reeves, R. R., Notarbartolo-Di-Sciara, G., Politi, E., Cañadas, A., Frantzis, A., & Mussi, B. (2003). Ecology, status and conservation of short-beaked common dolphins Delphinus delphis in the Mediterranean Sea: Common dolphins in the Mediterranean. Mammal Review, 33(3–4), 224–252. https://doi.org/10.1046/j.1365-2907.2003.00032.x
Birkun, A., Kuiken, T., Krivokhizhin, S., Haines, D. M., Osterhaus, A. D. M. E., van de Bildt, M. W. G., Joiris, C. R., & Siebert, U. (1999). Epizootic of morbilliviral disease in common dolphins (Delphinus delphis ponticus) from the Black Sea. Veterinary Record, 144(4), 85–92. https://doi.org/10.1136/vr.144.4.85
Biton Porsmoguer, S., Bənaru, D., Boudouresque, C., Dekeyser, I., Viricel, A., & Merchán, M. (2015). DNA evidence of the consumption of short-beaked common dolphin Delphinus delphis by the shortfin mako shark Isurus oxyrinchus. Marine Ecology Progress Series, 532, 177–183. https://doi.org/10.3354/meps11327
Braulik, G., Jefferson, T. A., & Bearzi, G. (2021). Delphinus delphis (amended version of 2021 assessment). The IUCN Red List of Threatened Species 2021: e.T134817215A199893039. https://doi.org/10.2305/IUCN.UK.2021-2.RLTS.T134817215A199893039.en
Brophy, J. T., Murphy, S., & Rogan, E. (2009). The diet and feeding ecology of the common dolphin (Delphinus delphis) in the northeast Atlantic. https://www.researchgate.net/publication/233385844
Cañadas, A., Donovan, G. P., Desportes, G., & Borchers, D. L. (2009). A short review of the distribution of short-beaked common dolphins (<i>Delphinus delphis</i>) in the central and eastern North Atlantic with an abundance estimate for part of this area. NAMMCO Scientific Publications, 7, 201–220. https://doi.org/10.7557/3.2714
Carwardine, M. (2020). Handbook of whales, dolphins, and porpoises of the world. Princeton University Press.
Castro, J., Faustino, C., Cid, A., Quirin, A., Matos, F. L., Rosa, R., & Pearson, H. C. (2022). Common dolphin (Delphinus delphis) fission–fusion dynamics in the south coast of Portugal. Behavioral Ecology and Sociobiology, 76(9). https://doi.org/10.1007/s00265-022-03235-0
Cockcroft, V. G., & Peddemorst, V. M. (1990). Seasonal distribution and density of common dolphins Delphinus delphis off the southeast coast of southern Africa. South African Journal of Marine Science, 9(1), 371–377.
Cunha, H. A., de Castro, R. L., Secchi, E. R., Crespo, E. A., Lailson-Brito, J., Azevedo, A. F., Lazoski, C., & Solé-Cava, A. M. (2015). Molecular and Morphological Differentiation of Common Dolphins (Delphinus sp.) in the Southwestern Atlantic: Testing the Two Species Hypothesis in Sympatry. PLoS ONE, 10(11), 1–15. https://doi.org/10.1371/journal.pone.0140251
Dabin, W., Cossais, F., & Ridoux, V. (2002). L’échouage en masse de Pleubian: une opportunité pour l’étude de la biologie et de la composition sociale du dauphin commun. Actes Du 4ème Séminaire Du Réseau National d’Echouages, Centre de Recherche Sur Les Mammifères Marins, 11.
Danil, K., & St. Leger, J. A. (2011). Seabird and Dolphin Mortality Associated with Underwater Detonation Exercises. Marine Technology Society Journal, 45(6), 89–95. https://doi.org/10.4031/MTSJ.45.6.5
Deaville, R. (2018). UK Cetacean Strandings Investigation Programme final contract report to Defra (MB0111 2011-2017).
Dool, T., & Bosker, T. (2022). Predicted microplastic uptake through trophic transfer by the short-beaked common dolphin (Delphinus delphis) and common bottlenose dolphin (Tursiops truncatus) in the Northeast Atlantic Ocean and Mediterranean Sea. Marine Pollution Bulletin, 180. https://doi.org/10.1016/J.MARPOLBUL.2022.113745
Evans, W. E. (1994). Common dolphin, white-bellied porpoise Delphinus delphis Linnaeus,1758. In S. H. Ridgway & R. Harrison (Eds.), Handbook of Marine Mammals (pp. 191–224).
Fearey, J., Elwen, S. H., James, B. S., & Gridley, T. (2019). Identification of potential signature whistles from free-ranging common dolphins (Delphinus delphis) in South Africa. Animal Cognition 2019 22:5, 22(5), 777–789. https://doi.org/10.1007/S10071-019-01274-1
Frantzis, A., & Herzing, D. L. (2002). Mixed-species associations of striped dolphins (Stenella coeruleoalba), short-beaked common dolphins (Delphinus delphis), and Risso’s dolphins (Grampus griseus) in the Gulf of Corinth (Greece, Mediterranean Sea). Aquatic Mammals, 28(2), 188–197. https://www.researchgate.net/publication/237457059
Genov, T. 2023. Delphinus delphis (Europe assessment). The IUCN Red List of Threatened Species 2023: e.T134817215A219009724. Accessed on 21 December 2023.
Hammond, P. S., Bearzi, G., Bjørge, A., Forney, K., Karczmarski, L., Kasuya, T., Perrin, W. F., Scott, M. D., Wang, J. Y., Wells, R. S., & Wilson, B. (2008). Delphinus delphis. In IUCN Red List of Threatened Species. https://doi.org/10.2305/IUCN.UK.2008.RLTS.T6336A12649851.en
Hammond, P. S., Lacey, C., Gilles, A., Viquerat, S., Börjesson, P., Herr, H., Macleod, K., Ridoux, V., Santos, M. B., Scheidat, M., Teilmann, J., Vingada, J., Øien, N., Gillespie, D., Leaper, R., Sveegaard, S., & Antonio Vázquez, J. (2017). Estimates of cetacean abundance in European Atlantic waters in summer 2016 from the SCANS-III aerial and shipboard surveys. https://library.wur.nl/WebQuery/wurpubs/fulltext/414756
Hammond, P. S., Macleod, K., Berggren, P., Borchers, D. L., Burt, L., Cañadas, A., Desportes, G., Donovan, G. P., Gilles, A., Gillespie, D., Gordon, J., Hiby, L., Kuklik, I., Leaper, R., Lehnert, K., Leopold, M., Lovell, P., Øien, N., Paxton, C. G. M., … Vázquez, J. A. (2013). Cetacean abundance and distribution in European Atlantic shelf waters to inform conservation and management. Biological Conservation, 164, 107–122. https://doi.org/10.1016/j.biocon.2013.04.010
Hansen, S. S. (2020). Vagrant Species in the NAMMCO Management Area. NAMMCO Marine Mammal Student Symposium, 5 February 2020.
Hayes, S. A., Josephson, E., Maze-Foley, K., Rosel, P. E., & Wallace, J. (2022). U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments 2021. https://repository.library.noaa.gov/view/noaa/45014
Henderson, E. E., Hildebrand, J. A., Smith, M. H., & Falcone, E. A. (2012). The behavioral context of common dolphin (Delphinus sp.) vocalizations. Marine Mammal Science, 28(3), 439–460. https://doi.org/10.1111/J.1748-7692.2011.00498.X
Hernandez-Gonzalez, A., Saavedra, C., Gago, J., Covelo, P., Santos, M. B., & Pierce, G. J. (2018). Microplastics in the stomach contents of common dolphin (Delphinus delphis) stranded on the Galician coasts (NW Spain, 2005–2010). Marine Pollution Bulletin, 137, 526–532. https://doi.org/10.1016/J.MARPOLBUL.2018.10.026
ICES. (2015). Report of the Working Group on Marine Mammal Ecology (WGMME). www.ices.dk
Jaber, J. R., Pérez, J., Arbelo, M., Zafra, R., & Fernández, A. (2006). Pathological and immunohistochemical study of gastrointestinal lesions in dolphins stranded in the Canary Islands. Veterinary Record, 159(13), 410–414. https://doi.org/10.1136/vr.159.13.410
Jepson, P. D., Deaville, R., Acevedo-Whitehouse, K., Barnett, J., Brownlow, A., Brownell Jr., R. L., Clare, F. C., Davison, N., Law, R. J., Loveridge, J., Macgregor, S. K., Morris, S., Murphy, S., Penrose, R., Perkins, M. W., Pinn, E., Seibel, H., Siebert, U., Sierra, E., … Fernández, A. (2013). What Caused the UK’s Largest Common Dolphin (Delphinus delphis) Mass Stranding Event? PLoS ONE, 8(4), e60953. https://doi.org/10.1371/journal.pone.0060953
Kasuya, T., & Perrin, W. F. (2017). Small cetaceans of Japan: Exploitation and biology. Small Cetaceans of Japan: Exploitation and Biology, 1–475. https://doi.org/10.1201/B21801/SMALL-CETACEANS-JAPAN-TOSHIO-KASUYA-WILLIAM-PERRIN
Mannocci, L., Dabin, W., Augeraud-Véron, E., Dupuy, J.-F., Barbraud, C., & Ridoux, V. (2012). Assessing the Impact of Bycatch on Dolphin Populations: The Case of the Common Dolphin in the Eastern North Atlantic. PLoS ONE, 7(2), e32615. https://doi.org/10.1371/journal.pone.0032615
Marçalo, A., Nicolau, L., Giménez, J., Ferreira, M., Santos, J., Araújo, H., Silva, A., Vingada, J., & Pierce, G. J. (2018). Feeding ecology of the common dolphin (Delphinus delphis) in Western Iberian waters: has the decline in sardine (Sardina pilchardus) affected dolphin diet? Marine Biology, 165(3). https://doi.org/10.1007/s00227-018-3285-3
Massett, N., O’Callaghan, S. A., Ocallaghan, S. A., Massett, N. S. A., & Massett, N. (2020). Short-Beaked Common Dolphins (Delphinus delphis) Observed Bow-Riding Basking Sharks (Cetorhinus maximus). Aquatic Mammals, 46(5), 461–465. https://doi.org/10.1578/AM.46.5.2020.461
Methion, S., & Díaz López, B. (2021). Spatial segregation and interspecific killing of common dolphins (Delphinus delphis) by bottlenose dolphins (Tursiops truncatus). Acta Ethologica, 24(2), 95–106. https://doi.org/10.1007/S10211-021-00363-0
Mintzer, V. J., Diniz, K., & Frazer, T. K. (2018). The use of aquatic mammals for bait in global fisheries. Frontiers in Marine Science, 5(JUN). https://doi.org/10.3389/FMARS.2018.00191
Mirimin, L., Westgate, A., Rogan, E., Rosel, P., Read, A., Coughlan, J., & Cross, T. (2009). Population structure of short-beaked common dolphins (Delphinus delphis) in the North Atlantic Ocean as revealed by mitochondrial and nuclear genetic markers. Marine Biology, 156(5), 821–834. https://doi.org/10.1007/S00227-008-1120-Y/TABLES/5
Moore, S. E., & Ridgway, H. (1995). Whistles produced by common dolphins from the Southern California Bight. Aquatic Mammals, 21(1), 55–55.
Morick, D., Davidovich, N., Zemah-Shamir, Z., Bigal, E., Rokney, A., Ron, M., Blum, S. E., Fleker, M., Soto, E., Heckman, T. I., Lau, S. C. K., Wosnick, N., Tchernov, D., & Scheinin, A. P. (2022). First Isolation and Characterization of Streptococcus agalactiae From a Stranded Wild Common Dolphin (Delphinus delphis). Frontiers in Marine Science, 9. https://doi.org/10.3389/fmars.2022.824071
Moura, A. E., Natoli, A., Rogan, E., & Hoelzel, A. R. (2013). Atypical panmixia in a European dolphin species (Delphinus delphis): Implications for the evolution of diversity across oceanic boundaries. Journal of Evolutionary Biology, 26(1), 63–75. https://doi.org/10.1111/JEB.12032
Murphy, S., Collet, A., & Rogan, E. (2005). Mating Strategy in the male common dolphin (Delphinus delphis): What gonadal analysis tells us. Journal of Mammalogy, 86(6), 1247–1258. https://doi.org/10.1644/1545-1542(2005)86[1247:MSITMC]2.0.CO;2
Murphy, S., Evans, P. G. H., Pinn, E., & Pierce, G. J. (2021). Conservation management of common dolphins: Lessons learned from the North-East Atlantic. Aquatic Conservation: Marine and Freshwater Ecosystems, 31(S1), 137–166. https://doi.org/10.1002/AQC.3212
Murphy, S., Herman, J. S., Pierce, G. J., Rogan, E., & Kitchener, A. C. (2006a). Taxonomic status and geographical cranial variation of common dolphins (Delphinus) in the Eastern North Atlantic. Marine Mammal Science, 22(3), 573–599. https://doi.org/10.1111/j.1748-7692.2006.00037.x
Murphy, S., Herman, J. S., Pierce, G. J., Rogan, E., & Kitchener, A. C. (2006b). Taxonomic status and geographical cranial variation of common dolphins (Delphinus) in the Eastern North Atlantic. Marine Mammal Science, 22(3), 573–599. https://doi.org/10.1111/j.1748-7692.2006.00037.x
Murphy, S., Winship, A., Dabin, W., Jepson, P. D., Deaville, R., Reid, R. J., Spurrier, C., Rogan, E., López, A., González, A. F., Read, F. L., Addink, M., Silva, M., Ridoux, V., Learmonth, J. A., Pierce, G. J., & Northridge, S. P. (2009). Importance of biological parameters in assessing the status of Delphinus delphis. Marine Ecology Progress Series, 388, 273–291. https://doi.org/10.3354/meps08129
Mussi, B., Vivaldi, C., Zucchini, A., Miragliuolo, A., & Pace, D. S. (2021). The decline of short-beaked common dolphin (Delphinus delphis) in the waters off the island of Ischia (Gulf of Naples, Italy). Aquatic Conservation: Marine and Freshwater Ecosystems, 31(S1), 87–100. https://doi.org/10.1002/AQC.3061
Natoli, A., Cañadas, A., Peddemors, V. M., Aguilar, A., Vaquero, C., Fernández-Piqueras, P., & Hoelzel, A. R. (2006). Phylogeography and alpha taxonomy of the common dolphin (Delphinus sp.). Journal of Evolutionary Biology, 19(3), 943–954. https://doi.org/10.1111/J.1420-9101.2005.01033.X
Natoli, A., Cañadas, A., Vaquero, C., Politi, E., Fernandez-Navarro, P., & Hoelzel, A. R. (2008). Conservation genetics of the short-beaked common dolphin (Delphinus delphis) in the Mediterranean Sea and in the eastern North Atlantic Ocean. Conservation Genetics, 9(6), 1479–1487. https://doi.org/10.1007/s10592-007-9481-1
NOAA Fisheries. (2012). Mass Stranding Event. https://media.fisheries.noaa.gov/dam-migration/capecod_mass_stranding_factsheet2012.pdf
Perrin, W. F. (2018). Common Dolphin: Delphinus delphis. In B. Würsig, J. G. M. Thewissen, & K. M. Kovacs (Eds.), Encyclopedia of Marine Mammals (pp. 205–209). Academic Press. https://doi.org/10.1016/B978-0-12-804327-1.00095-9
Perrin, W. F., & Reilly, S. B. (1984). Reproductive Parameters of Dolphins and Small Whales of the Family Deiphinidae. Report International Whaling Commission (SPECIAL ISSUE 6), 6, 97–133.
Pierce, G. J., Santos, M. B., Murphy, S., Learmonth, J. A., Zuur, A. F., Rogan, E., Bustamante, P., Caurant, F., Lahaye, V., Ridoux, V., Zegers, B. N., Mets, A., Addink, M., Smeenk, C., Jauniaux, T., Law, R. J., Dabin, W., López, A., Farré, J. M. A., … Boon, J. P. (2008). Bioaccumulation of persistent organic pollutants in female common dolphins (Delphinus delphis) and harbour porpoises (Phocoena phocoena) from western European seas: Geographical trends, causal factors and effects on reproduction and mortality. Environmental Pollution, 153, 401–415.
Pinela, A. M., Borrell, A., & Aguilar, A. (2011). Common dolphin morphotypes: Niche segregation or taxonomy? Journal of Zoology, 284(4), 239–247. https://doi.org/10.1111/j.1469-7998.2011.00798.x
Piroddi, C., Bearzi, G., Gonzalvo, J., & Christensen, V. (2011). From common to rare: The case of the Mediterranean common dolphin. Biological Conservation, 144(10), 2490–2498. https://doi.org/10.1016/J.BIOCON.2011.07.003
Pons-Bordas, C., Hazenberg, A., Hernandez-Gonzalez, A., Pool, R. v., Covelo, P., Sánchez-Hermosin, P., López, A., Saavedra, C., Fraija-Fernández, N., Fernández, M., & Aznar, F. J. (2020). Recent increase of ulcerative lesions caused by Anisakis spp. In cetaceans from the north-east Atlantic. Journal of Helminthology. https://doi.org/10.1017/S0022149X20000115
Pusineri, C., Magnin, V., Meynier, L., Spitz, J., Hassani, S., & Ridoux, V. (2007). Food and feeding ecology of the common dolphin (Delphinus delphis) in the oceanic Northeast Atlantic and comparison with its diet in neritic areas. Marine Mammal Science, 23(1), 30–47. https://doi.org/10.1111/J.1748-7692.2006.00088.X
Reboredo-Fernández, A., Gómez-Couso, H., Martínez-Cedeira, J. A., Cacciò, S. M., & Ares-Mazás, E. (2014). Detection and molecular characterization of Giardia and Cryptosporidium in common dolphins (Delphinus delphis) stranded along the Galician coast (Northwest Spain). Veterinary Parasitology, 202(3–4), 132–137. https://doi.org/10.1016/j.vetpar.2014.03.018
Reidarson, T. H., McBain, J., House, C., King, D. P., Stott, J. L., Krafft, A., Taubenberger, J. K., Heyning, J., & Lipscomb, T. P. (1998). Morbillivirus infection in stranded common dolphins from the pacific ocean. Journal of Wildlife Diseases, 34(4), 771–776. https://doi.org/10.7589/0090-3558-34.4.771
Rogan, E., Breen, P., Mackey, M., Cañadas, A., Scheidat, M., Geelhoed, S., & Jessopp, M. (2018). Aerial Surveys of Cetaceans and Seabirds in Irish waters: Occurrence, distribution and abundance in 2015-2017. https://secure.dccae.gov.ie/downloads/SDCU_DOWNLOAD/ObSERVE_Aerial_Report.pdf
Simonis, A. E., Roch, M. A., Bailey, B., Barlow, J., Clemesha, R. E. S., Iacobellis, S., Hildebrand, J. A., & Baumann-Pickering, S. (2017). Lunar cycles affect common dolphin Delphinus delphis foraging in the Southern California Bight. Marine Ecology Progress Series, 577, 221–235. https://doi.org/10.3354/meps12247
Soldevilla, M. S., Henderson, E. E., & Campbell, G. S. (2008). Classification of Risso’s and Pacific white-sided dolphins using spectral properties of echolocation clicks. Citation: The Journal of the Acoustical Society of America, 124, 609. https://doi.org/10.1121/1.2932059
Souter, R., Chaber, A. L., Lee, K., Machado, A., Lam, J., & Woolford, L. (2021). Fatal Streptococcus iniae Infection in a Juvenile Free-Ranging Short-Beaked Common Dolphin (Delphinus delphis). Animals, 11(11). https://doi.org/10.3390/ani11113123
Thompson, F. N., Abraham, E. R., & Berkenbusch, K. (2013). Common Dolphin (Delphinus delphis) Bycatch in New Zealand Commercial Trawl Fisheries. PLoS ONE, 8(5), e64438. https://doi.org/10.1371/journal.pone.0064438
Viricel, A., Strand, A. E., Rosel, P. E., Ridoux, V., & Garcia, P. (2008). Insights on common dolphin (Delphinus delphis) social organization from genetic analysis of a mass-stranded pod. Behavioral Ecology and Sociobiology, 63(2), 173–185. https://doi.org/10.1007/s00265-008-0648-7
Würsig, B. (2018). Bow-Riding. In B. Würsig, J. G. M. Thewissen, & K. M. Kovacs (Eds.), Encyclopedia of Marine Mammals (pp. 135–137). Academic Press. https://doi.org/10.1016/B978-0-12-804327-1.00076-5
Young, D. D., & Cockcroft, V. G. (1994). Diet of common dolphins Delphinus delphis off the south‐east coast of southern Africa: opportunism or specialization? Journal of Zoology, 234(1), 41–53. https://doi.org/10.1111/j.1469-7998.1994.tb06055.x
Zanardo, N., Bilgmann, K., Parra, G. J., & Möller, L. M. (2016). Socio-genetic structure of short-beaked common dolphins in southern Australia. Journal of Zoology, 299(2), 89–97. https://doi.org/10.1111/jzo.12330
Zemsky, V. A. (1996). History of the Russian fishery of dolphins in the Black Sea. In B. Öztürk (Ed.), Proceedings of the First International Symposium on the Marine Mammals of the Black Sea (pp. 46–48).